تنظیم سنتز کلسترول
تنظیم سنتز کلسترول - آنزیم اصلی آن (HMG-CoA ردوکتاز) به روش های مختلفی انجام می شود.
فسفوریلاسیون / دفسفوریلاسیون ردوکتاز HMG. با افزایش نسبت انسولین / گلوکاگون ، این آنزیم فسفریله می شود و به حالت فعال منتقل می شود. عمل انسولین از طریق 2 آنزیم انجام می شود.
HMG-CoA ردوکتاز کیناز فسفاتاز ، که کیناز را به حالت دفسفریله غیرفعال تبدیل می کند:
فسفوتاز HMG-CoA ردوکتاز با تبدیل آن به حالت فعال دفسفریله شده. نتیجه این واکنشها تشکیل یک فرم فعال دفسفریله شده از HMG-CoA ردوکتاز است.
در نتیجه ، در دوره جذب ، کلسترول افزایش می یابد. در این دوره ، در دسترس بودن بستر اولیه برای سنتز کلسترول - استیل - CoA افزایش می یابد (در نتیجه خوردن مواد غذایی حاوی کربوهیدرات ها و چربی ها ، از آنجا که استیل CoA استیل در هنگام تجزیه گلوکز و اسیدهای چرب تشکیل می شود).
در حالت پس از جذب ، گلوکاگون از طریق پروتئین ژناز A فسفوریلاسیون HMG - CoA - ردوکتاز را تحریک می کند ، و آن را به حالت غیرفعال تبدیل می کند. این عمل با این واقعیت تقویت می شود که در همان زمان گلوکاگون باعث تحریک فسفوریلاسیون و غیرفعال کردن فسفوتاز ردوکتاز HMG-CoA می شود ، بنابراین ردوکتاز HMG-CoA را در حالت غیرفعال فسفریله نگه می دارد. در نتیجه ، سنتز کلسترول در دوره پس از جذب و در طول روزه دار با سنتز درون زا مهار می شود. اگر میزان کلسترول موجود در مواد غذایی به 2٪ برسد ، در نتیجه سنتز کلسترول درون زا به شدت کاهش می یابد. اما قطع کامل سنتز کلسترول رخ نمی دهد.
میزان مهار بیوسنتز کلسترول تحت تأثیر کلسترول ناشی از غذا در افراد متفاوت است. این نشان دهنده فردیت فرآیندهای تشکیل کلسترول است. با کاهش شدت سنتز کلسترول می توان غلظت آن را در خون کاهش داد.
اگر تعادل بین دریافت کلسترول با مواد غذایی و سنتز آن در بدن از یک طرف و دفع اسیدهای صفراوی و کلسترول از طرف دیگر شکسته شود ، غلظت کلسترول در بافت ها و خون تغییر می کند. جدی ترین عواقب با افزایش غلظت کلسترول خون (هایپر کلسترول خون) همراه است ، در حالی که احتمال ابتلا به آترواسکلروز و کللیتیزیس افزایش می یابد.
هایپرکلسترولمی خانوادگی (HCS) - این شکل بسیار متداول است - در حدود 200 بیمار در حدود 1 بیمار. نقص ارثی در HCS یک تخلف در جذب LDL توسط سلول ها است و بنابراین کاهش میزان کاتابولیسم LDL است. در نتیجه ، غلظت LDL در خون بالا می رود ، همچنین کلسترول از آنجا که مقدار زیادی از آن در LDL وجود دارد. بنابراین ، با HCS ، رسوب کلسترول در بافت ها ، به ویژه در پوست (زانتیوما) ، در دیواره شریان ها مشخص است.
مهار سنتز ردوکتاز HMG-CoA
محصول نهایی مسیر متابولیک کلسترول. این میزان رونویسی از ژن HMG-CoA ردوکتاز را کاهش می دهد ، بنابراین سنتز خاص خود را مهار می کند. کبد به طور فعال اسیدهای صفراوی را از کلسترول تولید می کند ، بنابراین اسیدهای صفراوی فعالیت ژن HMG-CoA ردوکتاز را مهار می کنند. از آنجا که ردوکتاز HMG-CoA پس از سنتز حدود 3 وجود دارد ، مهار سنتز این آنزیم کلسترول یک تنظیم مؤثر است.
آنچه را که می خواستید پیدا نکردید؟ استفاده از جستجو:
تبادل کلسترول استر
صندوق کلسترول حاوی کلسترول و استرهای کلسترول آزاد است ، که هم در سلول ها و هم در لیپوپروتئین های خون یافت می شوند.
قسمت دوم متابولیسم و انرژی
در سلولها ، استری شدن کلسترول با عملکرد اسید-CoA-کلسترول- acyltransferase (AChAT) اتفاق می افتد:
Acyl-CoA + کلسترول - * HS-KoA + کلسترول
در سلول های انسانی ، لینوللی کلسترول به طور عمده تشکیل می شود. برخلاف کلسترول آزاد ، استرهای آن در غشای سلولی به مقدار بسیار کمی موجود است و به طور عمده در سیتوزول به عنوان بخشی از قطرات لیپید یافت می شود. تشکیل استرها از یک طرف می تواند مکانیسمی برای از بین بردن کلسترول اضافی از غشاها در نظر گرفته شود و از سوی دیگر به عنوان مکانیسم ذخیره کلسترول در سلول در نظر گرفته شود. بسیج منابع با مشارکت آنزیم های استراز که هیدرولیز استرهای کلسترول هیدرولیز می شوند رخ می دهد:
اسید چرب + کلسترول
سنتز و هیدرولیز استرها در بسیاری از سلول ها رخ می دهد ، اما به ویژه در سلول های قشر آدرنال فعال است: در این سلول ها تا 80٪ کل کلسترول توسط استرها نشان داده می شوند ، در حالی که در سلولهای دیگر معمولاً کمتر از 20٪ است.
تشکیل استرها در لیپونروتئین های خون با مشارکت لسیتین کلسترول آسیل ترانسفراز (LHAT) صورت می گیرد ، که انتقال باقی مانده آسیلی را از موقعیت i i لسیتین به کلسترول کاتالیز می کند (شکل 10.31) LHAT در کبد تشکیل می شود ، ترشح شده در خون و به لیپوپروتئین ها متصل می شود. میزان استری سازی برای لیپوپروتئین های مختلف به طور قابل توجهی متفاوت است و به حضور آپولیپوپروتئین هایی که LHAT را فعال می کنند (عمدتا آپو-AT و همچنین CI) یا مهار کننده (C-II) این آنزیم است.فعال ترین LHAT در HDL ، که در آن apo-AT باید olee 2/3 از همه پروتئین. بزرگترین افتخارات کولی استرهای اسیدهای اولئیک و لینولئیک تشکیل شده است. سایر لیپوپروتئینها استر تشکیل با سرعت کند تر نسبت به HDL رخ می دهد.
شکل 10.31 تشکیل استرهای کلسترول تحت عمل LHAT
LHAT در لایه سطحی HDL بومی سازی شده است و از کلسترول موجود در یک لایه فسفولیپید به عنوان بستر استفاده می کند. استرهای کلسترول که در اینجا تشکیل شده اند ، به دلیل آبگریزگی کامل ، ضعیف در آن نگهداری می شوند
فصل 10. متابولیسم و عملکرد لیپیدها
فسفولیپید تک لایه و غوطه ور در هسته چربی لیپوپروتئین. در عین حال ، مکانی برای کلسترول در تک لایه فسفولیپید آزاد می شود که می تواند از غشای سلولی یا سایر لیپوپروتئین ها با کلسترول پر شود. بنابراین ، HDL به عنوان یک نتیجه از عملکرد LHAT به عنوان یک تله کلسترول به نظر می رسد.
سنتز اسیدهای صفراوی
در کبد ، بخشی از کلسترول به اسیدهای صفراوی تبدیل می شود. اسیدهای گالیک می توانند به عنوان مشتقات اسید کولانیک در نظر گرفته شوند (شکل 10.32).
اسید کلانونیک به همین ترتیب در بدن شکل نمی گیرد. در سلولهای کبدی ، کلسترول مستقیماً اسیدهای فنوتیکولیکولیکولیکولی و کلولیکی را تولید می کند - اسیدهای صفراوی اولیه (شکل 10.33 ، همچنین به شکل 10.12 مراجعه کنید).
بیوسنتز کلسترول
بیوسنتز کلسترول در شبکه آندوپلاسمی رخ می دهد. منبع تمام اتمهای کربن موجود در این مولکول استیل-SCoA است که در اینجا از میتوکندریا به عنوان بخشی از سیترات و همچنین در سنتز اسیدهای چرب حاصل می شود. بیوسنتز کلسترول 18 مولکول ATP و 13 مولکول NADPH مصرف می کند.
تشکیل کلسترول در بیش از 30 واکنش رخ می دهد که می توانند در چند مرحله گروه بندی شوند.
1. سنتز اسید ماوالونیک.
دو واکنش سنتز اول همزمان با واکنش کتوژنز است ، اما پس از سنتز 3-هیدروکسی-3-متیل گلوتاریل-ScoA ، آنزیم وارد می شود. هیدروکسی متیل گلوتاریل-ScoA ردوکتاز (HMG-SCOA ردوکتاز) ، تشکیل اسید میاوونیک.
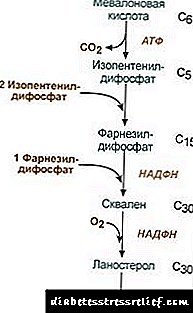 |
طرح واکنش سنتز کلسترول2. سنتز ایزوپنتنیل دیفسفات. در این مرحله ، سه باقیمانده فسفات به اسید میوونونیک متصل می شوند ، سپس آن را دکربوکسیله و دهیدروژن می شود. 3. پس از ترکیب سه مولکول ایزوپنتنیل دیفسفات ، فارنسیل دیفسفات سنتز می شود. 4- سنتز اسكالن زمانی متصل می شود كه دو باقیمانده فارنسیل دیفسفات به هم وصل شوند. 5- بعد از واكنش هاي پيچيده ، اسكالن خطي به لانسترول چرخه مي دهد. 6. از بین بردن گروه های متیل اضافی ، ترمیم و ایزومریزاسیون مولکول منجر به بروز کلسترول می شود. تنظیم فعالیت ردوکتاز هیدروکسی متیل گلوتاریل-S-CoA3. میزان بیوسنتز کلسترول نیز به غلظت آن بستگی دارد پروتئین حامل خاصتأمین اتصال و انتقال متابولیتهای سنتز واسطه آبگریز می توانید نظر خود را بپرسید یا بگذارید. نکته اصلی تنظیم ، واکنش تشکیل اسید میوونیک اسید است. 1. مقررات آلوسرتیک. کلسترول ، و در کبد - و اسیدهای صفراوی مانع ردوکتاز HMG-CoA می شوند. 2. سرکوب سنتز کلسترول HMG-CoA ردوکتاز. 3. تنظیم توسط فسفوریلاسیون-دفسفوریلاسیون HMG-CoA ردوکتاز ، فرم غیر فسفریله فعال. گلوکاگون باعث غیرفعال شدن می شود و انسولین باعث فعال شدن از طریق آبشار پیچیده ای از واکنشها می شود. بنابراین ، میزان سنتز کلسترول با تغییر در حالت های جاذب و جذب جذب می شود. 4- میزان سنتز ردوکتاز HMG-CoA در کبد در معرض نوسانات روزانه روشن است: حداکثر در نیمه شب و حداقل در صبح. تبادل کلسترول استر در سلولها استری شدن کلسترول در هنگام مواجهه رخ می دهد acyl-CoA- کلسترول- acyltransferase (AHAT): Acyl-CoA + کلسترول ® NS-CoA + کلسترول در سلولها ، به طور عمده لینولای کلسترول تشکیل می شود. استرها به عنوان بخشی از قطرات لیپیدی به طور عمده در سیتوزول یافت می شوند. تشکیل استرها از یک طرف می تواند مکانیسمی برای از بین بردن کلسترول اضافی از غشاها در نظر گرفته شود و از طرف دیگر به عنوان مکانیزمی برای ذخیره کلسترول در سلول در نظر گرفته شود. بسیج ذخایر با مشارکت آنزیم ها اتفاق می افتد استرازهیدرولیز کردن استرهای کلسترول: اسید کلسترول + N2O Ac اسیدهای چرب + کلسترول سنتز و هیدرولیز استرها به ویژه در سلولهای قشر آدرنال فعال است. در لیپوپروتئین های خون تشکیل استر با مشارکت لسیتین-کلسترول- آسییل ترانسفراز (LHAT) ، کاتالیزور انتقال باقیمانده acyl از لسیتین به کلسترول. LHAT در کبد تشکیل می شود ، در جریان خون ترشح می شود و به لیپوپروتئین ها وصل می شود. LHAT فعال ترین در HDL ، جایی که در لایه سطح بومی سازی شده است. استرهای کلسترول ایجاد شده در اینجا ، آبگریز بوده و در هسته لیپید غوطه ور می شوند. در تک لایه فسفولیپید ، فضای خالی برای کلسترول وجود دارد که می تواند با کلسترول از غشای سلولی یا از دیگر لیپوپروتئین ها پر شود. بنابراین ، HDL به عنوان یک نتیجه از عملکرد LHAT به عنوان یک تله کلسترول به نظر می رسد. سنتز اسیدهای صفراوی در کبد ، بخشی از کلسترول به اسیدهای صفراوی تبدیل می شود. اسیدهای صفراوی را می توان مشتقات اسید کلانیک دانست. اسید کلانونیک به همین ترتیب در بدن شکل نمی گیرد. در سلولهای کبدی از کلسترول ، اسیدهای صفراوی اولیه تشکیل می شوند - chenodeoxycholic و چولی. پس از ترشح صفرا به روده تحت عمل آنزیم های فلور روده ، اسیدهای صفراوی ثانویه از آنها تشکیل می شود - سنگ سنگی و دیوکسیکولیک. آنها از روده ها جذب می شوند ، با خون ورید پرتال وارد کبد و سپس به صفرا می شوند. صفرا به طور عمده حاوی اسیدهای صفراوی کونژوگه ، یعنی ترکیبات آنها با گلیسین یا تورین است. غلظت اسیدهای صفراوی در صفرا حدود 1٪ است. در قسمت اصلی اسیدهای صفراوی نقش دارد گردش خون کبدی.بخش کمی از اسیدهای صفراوی - حدود 0.5 گرم در روز - در مدفوع دفع می شود. این با سنتز اسیدهای صفراوی جدید در کبد جبران می شود ، صندوق اسید صفراوی در حدود 10 روز به روز می شود. کلسترول نیز به طور عمده از طریق روده ها دفع می شود. با غذا و از کبد به عنوان بخشی از صفرا وارد روده ها می شود. کلسترول جذب شده در خون حاوی کسری از صفرا است (کلسترول درون زادر کبد سنتز می شود و کسری که از مواد غذایی حاصل می شود (کلسترول اگزوژن) حذف کلسترول از بافت ها با اکسیداسیون آن به اسیدهای صفراوی در کبد صورت می گیرد و به دنبال آن دفع آنها با مدفوع (تقریباً 0.5 گرم در روز) و با دفع کلسترول بدون تغییر (همچنین با مدفوع) صورت می گیرد. در حالت ثابت: (کلسترولپایان + کلسترولسابق) - (کلسترولطرد + اسیدهای صفراویطرد) = 0 اگر این تعادل مختل شود ، غلظت کلسترول در بافت ها و خون تغییر می کند. افزایش کلسترول خون - فشار خون بالا. این احتمال آترواسکلروز و بیماری کیسه صفرا را افزایش می دهد. مقررات تبادل LIPID سوخت و ساز چربی توسط سیستم عصبی مرکزی تنظیم می شود. ماندگار استرس عاطفی منفیافزایش انتشار کاتکول آمین ها در جریان خون باعث کاهش قابل توجه وزن می شود. عمل گلوکاگون در سیستم لیپولیتیک شبیه عمل کاتکول آمین ها است. آدرنالین و نوراپی نفرین فعالیت لیپاز بافتی و میزان لیپولیز در بافت چربی را افزایش می دهد ، در نتیجه ، محتوای اسیدهای چرب در پلاسما خون افزایش می یابد. انسولین اثر متضاد آدرنالین و گلوکاگون بر لیپولیز و بسیج اسیدهای چرب دارد. هورمون رشد تحریک لیپولیز ، القاء سنتز acenylate cyclase. عملکرد هیپوفیز باعث رسوب چربی در بدن (چاقی هیپوفیز) می شود. تیروکسین ، هورمونهای جنسیهمچنین بر متابولیسم چربی تأثیر می گذارد. از بین بردن غدد جنسی در حیوانات باعث رسوب بیش از حد چربی می شود. دستگاه های متابولیک لیپید کلسترول یک استروئید خاص برای ارگانیسم های حیوانات است. محل اصلی شکل گیری آن در بدن انسان کبد است ، که در آن 50٪ کلسترول سنتز می شود ، 15-20٪ در روده کوچک تشکیل می شود ، بقیه در پوست ، قشر آدرنال و غدد جنسی سنتز می شوند. منابع تشکیل صندوق کلسترول و راه های هزینه آن در شکل 22.1 ارائه شده است. شکل 22.1. تشکیل و توزیع کلسترول در بدن. کلسترول بدن انسان (مقدار کل حدود 140 گرم) به طور شرطی می تواند به سه استخر تقسیم شود: 30 گرم) ، به سرعت در حال تبادل ، متشکل از کلسترول دیواره روده ، پلاسمای خون ، کبد و سایر اندامهای پارانشیمی است ، تجدید در 30 روز (1 گرم در روز) اتفاق می افتد ، 50 گرم) ، به آرامی در حال تبادل کلسترول ارگان ها و بافت ها ، 60 گرم) ، یک کلسترول بسیار به آرامی رد و بدل شده از نخاع و مغز ، بافت همبند ، میزان بروزرسانی در طول سالها محاسبه می شود. سنتز کلسترول در سیتوزول سلول ها اتفاق می افتد. این یکی از طولانی ترین مسیرهای متابولیکی در بدن انسان است. در 3 مرحله ادامه می یابد: اولین مرحله با تشکیل اسید میوالونیک ، دوم با تشکیل اسکالن (ساختار هیدروکربن خطی متشکل از 30 اتم کربن) به پایان می رسد. در مرحله سوم ، اسکالن به یک مولکول لانوسترول تبدیل می شود ، سپس 20 واکنش پی در پی وجود دارد که لانوسترول را به کلسترول تبدیل می کند. در برخی از بافت ها ، گروه هیدروکسیل کلسترول استر را تشکیل می دهد و استرها را تشکیل می دهند. این واکنش توسط آنزیم داخل سلولی AHAT کاتالیز می شود (acylCoA: کلسترول آسیل ترانسفراز). واکنش استری سازی نیز در HDL ، جایی که آنزیم LHAT (لسیتین: کلسترول آسیل ترانسفراز) در آن قرار دارد ، در خون رخ می دهد. استرهای کلسترول شکلی است که در آن با خون منتقل می شود یا در سلول ها رسوب می شود. در خون ، حدود 75٪ کلسترول به شکل استر است. سنتز کلسترول با تأثیر بر فعالیت و میزان آنزیم کلیدی فرآیند تنظیم می شود - 3-هیدروکسی-3-متیلگلوتاریل-CoA ردوکتاز (HMG-CoA ردوکتاز). این به دو روش حاصل می شود: 1. فسفوریلاسیون / دفسفوریلاسیون ردوکتاز HMG-CoA. انسولین دفسفوریلاسیون HMG-CoA ردوکتاز را تحریک می کند ، از این طریق آن را به حالت فعال تبدیل می کند. بنابراین ، در دوره جذب ، سنتز کلسترول افزایش می یابد. در این دوره ، در دسترس بودن بستر شروع برای سنتز ، استیل CoA نیز افزایش می یابد. گلوکاگون تأثیر متضاد دارد: از طریق پروتئین کیناز A ، فسفوریلاسیون HMG-CoA ردوکتاز را تحریک می کند ، و آن را به حالت غیرفعال تبدیل می کند. در نتیجه ، سنتز کلسترول در دوره پس از جذب و در طول روزه دار مهار می شود. 2. مهار سنتز ردوکتاز HMG-CoA.کلسترول (محصول نهایی مسیر متابولیک) میزان رونویسی ژن HMG-CoA ردوکتاز را کاهش می دهد ، بنابراین سنتز خود را مهار می کند و اسیدهای صفراوی نیز باعث ایجاد یک اثر مشابه می شوند. انتقال کلسترول خون به عنوان بخشی از دارو انجام می شود. LP ها کلسترول برون زا را در بافت ها فراهم می کنند ، جریان آن بین اعضای بدن و دفع بدن را تعیین می کنند. کلسترول اگزوژن به عنوان بخشی از ChM باقیمانده به کبد تحویل داده می شود. در آنجا ، همراه با کلسترول درون زا سنتز ، یک صندوق مشترک را تشکیل می دهد. در سلولهای کبدی TAG و کلسترول در VLDL بسته بندی می شوند و به این شکل در خون ترشح می شوند. در خون ، VLDL تحت عمل LP-lipase ، که TAG را به گلیسرول و اسیدهای چرب هیدرولیز می کند ، ابتدا به LSPP تبدیل می شود ، و سپس به LDL که حاوی 55 درصد کلسترول و استرهای آن است. LDL اصلی ترین نوع حمل و نقل کلسترول است که در آن به بافت ها تحویل داده می شود (70٪ کلسترول و استرهای آن در خون بخشی از LDL هستند). LDL از خون وارد کبد (تا 75٪) و سایر بافتهایی که دارای گیرنده های LDL در سطح آنها هستند می شود. اگر مقدار کلسترول وارد شده به سلول از نیاز آن فراتر رود ، سنتز گیرنده های LDL سرکوب می شود که باعث کاهش جریان کلسترول خون می شود. برعکس ، با کاهش غلظت کلسترول آزاد در سلول ، سنتز گیرنده فعال می شود. هورمون ها در تنظیم سنتز گیرنده LDL دخیل هستند: انسولین ، تریودوتیرونین و هورمون های جنسی شکل گیری گیرنده ها را افزایش می دهند و گلوکوکورتیکوئیدها کاهش می یابد. در به اصطلاح "انتقال کلسترول بازگشت" ، یعنی مسیری که بازگشت کلسترول به کبد را تضمین می کند ، HDL نقش اصلی را ایفا می کند. آنها در کبد به شکل پیش سازهای نابالغ ساخته می شوند که عملا حاوی کلسترول و TAG نیستند. پیش سازهای HDL در خون توسط کلسترول اشباع شده و آن را از سایر LP ها و غشاهای سلولی دریافت می کنند. انتقال کلسترول به HDL شامل آنزیم LHAT واقع در سطح آنها است. این آنزیم باقیمانده اسید چرب را از فسفاتیدیل کلولین (لسیتین) به کلسترول متصل می کند. در نتیجه ، یک مولکول آبگریز کلسترول استر تشکیل می شود ، که درون HDL حرکت می کند. بنابراین ، HDL مست نیست ، غنی از کلسترول ، تبدیل به HDL 3 - ذرات بالغ و بزرگتر. HDL 3 تبادل استرهای کلسترول برای TAG موجود در VLDL و STD با مشارکت یک پروتئین خاص که استرهای کلسترول را بین لیپوپروتئین ها منتقل می کند. در این حالت ، HDL 3 تبدیل به HDL2 ، به دلیل تجمع TAG اندازه آن افزایش می یابد. VLDL و STDL تحت تأثیر LP-lipase به LDL تبدیل می شوند که عمدتاً کلسترول را به کبد منتقل می کنند. بخش کوچکی از کلسترول به کبد HDL2 و HDL منتقل می شود. سنتز اسیدهای صفراوی. در کبد ، 500-700 میلی گرم اسیدهای صفراوی در روز از کلسترول ساخته می شود. تشکیل آنها شامل واکنشهای معرفی گروههای هیدروکسیل با مشارکت هیدروکسیلازها و واکنش اکسیداسیون جزئی زنجیره جانبی کلسترول است (شکل 22.2): شکل 22.2 طرح تشکیل اسیدهای صفراوی. اولین واکنش سنتز - تشکیل 7-هیدروکسی کلسترول - نظارتی است. فعالیت آنزیمی که این واکنش را کاتالیز می کند با محصول نهایی مسیر ، اسیدهای صفراوی مهار می شود. مکانیسم تنظیم دیگر فسفوریلاسیون / دفسفوریلاسیون آنزیم است (فرم فسفریله شده 7-هیدروکسیلاز فعال است). تنظیم با تغییر مقدار آنزیم نیز امکان پذیر است: کلسترول باعث رونویسی از ژن 7-هیدروکسیلاز و سرکوب اسیدهای صفراوی می شود. هورمونهای تیروئید سنتز 7-هیدروکسیلاز و سرکوب استروژن را القا می کنند. چنین تأثیر استروژن در سنتز اسیدهای صفراوی توضیح می دهد که چرا بیماری سنگ کیسه صفرا در زنان 3-4 برابر بیشتر از مردان رخ می دهد. اسیدهای کولیک و فنای سنتوکسیکولیک که از کلسترول تشکیل شده اند "اسیدهای صفراوی اولیه" نامیده می شوند. بخش عمده ای از این اسیدها تحت آزمایش قرار می گیرند - افزودن مولکول های گلیسین یا تورین به گروه کربوکسیل اسید صفراوی. کونژوگه با تشکیل شکل فعال اسیدهای صفراوی - مشتقات CoA ، سپس تورین یا گلیسین به هم متصل می شوند و در نتیجه 4 نوع ترکیب مزدوج تشکیل می شود: تائوروکولیک و تائوروئندوکسیکولیک ، گلیکوکولیک و اسیدهای گلیکوئیندوکسیکولیک. آنها امولسیفایر قابل ملاحظه ای قوی تر از اسیدهای صفراوی اصلی هستند. ترکیبات گلیسین 3 برابر بیشتر از تائورین تشکیل می شوند ، زیرا مقدار تورین در بدن محدود است. در روده ، مقدار کمی از ترکیبات اسیدهای صفراوی اولیه تحت اثر آنزیم های باکتریایی به اسیدهای صفراوی ثانویه تبدیل می شوند. دیوکسیکولیک اسید ، که از کلسترول تشکیل شده است ، و لیتوچولیک ، که از دیوکسیکولیک تشکیل شده است ، محلول کمتری و آهسته تر در روده ها جذب می شوند. حدود 95٪ اسیدهای صفراوی که وارد روده می شوند از طریق ورید پورتال به کبد برمی گردند ، سپس دوباره در صفرا ترشح می شوند و در امولسیون چربی ها مجدداً مورد استفاده قرار می گیرند. به این مسیر اسیدهای صفراوی گردش خون انتروهپاتی گفته می شود. با مدفوع ، اسیدهای صفراوی ثانویه بیشتر حذف می شوند. بیماری سنگ کیسه صفرا (کللیتیاسیس) یک فرآیند آسیب شناختی است که در آن سنگ ها در کیسه صفرا تشکیل می شود ، که اساس آن کلسترول است. ترشح کلسترول به داخل صفرا باید با انتشار متناسب اسیدهای صفراوی و فسفولیپیدها باشد که مولکولهای کلسترول آبگریز را در حالت میسل نگه می دارد. دلایلی که منجر به تغییر در نسبت اسیدهای صفراوی و کلسترول در صفرا می شود عبارتند از: مواد غذایی سرشار از کلسترول ، غذای پر کالری ، رکود صفرا در کیسه صفرا ، اختلال در گردش خون روده ای ، اختلال در سنتز اسیدهای صفراوی ، عفونت کیسه صفرا. در بیشتر بیماران مبتلا به كللیتیتاز ، سنتز كلسترول افزایش یافته و سنتز اسیدهای صفراوی از آن كند می شود و این منجر به عدم تناسب در تعداد كلسترول و اسیدهای صفراوی ترشح شده در صفرا می شود. در نتیجه ، کلسترول در کیسه صفرا شروع به رسوب می کند و یک رسوب چسبناک تشکیل می دهد که به تدریج سخت می شود. بعضی اوقات با بیلی روبین ، پروتئین ها و نمک های کلسیم آغشته می شود. سنگ ها فقط می توانند از کلسترول (سنگ های کلسترول) یا ترکیبی از کلسترول ، بیلی روبین ، پروتئین ها و کلسیم تشکیل شوند. سنگ های کلسترول معمولاً سفید هستند و سنگ های مختلط در سایه های مختلف قهوه ای هستند. در مرحله اولیه شکل گیری سنگ ، از اسید فنولی فنولیک اسید می توان به عنوان دارو استفاده کرد. هنگامی که در کیسه صفرا قرار دارد ، به تدریج سنگ های کلسترول را حل می کند ، اما این یک روند کند است که چندین ماه طول می کشد. آترواسکلروز یک آسیب شناسی است که با ظاهر شدن پلاکهای آتروژنیک روی سطح داخلی دیواره عروقی مشخص می شود. یکی از دلایل اصلی ایجاد چنین پاتولوژی ، نقض تعادل بین مصرف کلسترول از مواد غذایی ، سنتز آن و دفع آن از بدن است. بیماران مبتلا به آترواسکلروز غلظت LDL و VLDL را افزایش داده اند. بین غلظت HDL و احتمال بروز آترواسکلروز رابطه معکوس وجود دارد. این با مفهوم عملکرد LDL به عنوان حامل کلسترول در بافت ها و HDL از بافت سازگار است. "شرط اساسی" متابولیک برای ایجاد آترواسکلروز ، پرکلسترول خون است. (کلسترول بالا در خون). هایپرکلسترولمی ایجاد می شود: 1. به دلیل مصرف زیاد کلسترول ، کربوهیدرات ها و چربی ها ، 2- پیش بینی ژنتیکی متشکل از نقص های ارثی در ساختار گیرنده های LDL یا apoB-100 ، و همچنین در افزایش سنتز یا ترشح apoB-100 (در مورد هایپرلیپیدمی ترکیبی خانوادگی ، که در آن غلظت خون و کلسترول و TAG بالا می رود). نقش مهمی در مکانیسم های توسعه آترواسکلروز با اصلاح داروی بازی می شود. تغییر در ساختار طبیعی لیپیدها و پروتئین ها در LDL باعث می شود آنها برای بدن بیگانه باشند و بنابراین برای گرفتن فاگوسیت ها در دسترس تر هستند. اصلاح دارو می تواند توسط مکانیسم های مختلفی رخ دهد: 1. گلیکوزیلاسیون پروتئین که با افزایش غلظت گلوکز در خون رخ می دهد ، 2. اصلاح پراکسید ، منجر به تغییر در لیپیدها در لیپوپروتئین ها و ساختار apoB-100 ، 3. تشکیل مجتمع های خود ایمنی LP-آنتی بادی (داروهای تغییر یافته می توانند باعث ایجاد آنتی بادی ها شوند). LDL اصلاح شده توسط ماکروفاژها جذب می شود. این فرآیند با میزان کلسترول جذب شده تنظیم نمی شود ، همانطور که در مورد ورود آن به سلول ها از طریق گیرنده های خاص ، بنابراین ماکروفاژها با کلسترول بارگیری می شوند و به "سلول های فوم" تبدیل می شوند که به فضای subendothelial نفوذ می کنند. این منجر به ایجاد لکه های چربی یا نوارهایی در دیواره رگ های خونی می شود. در این مرحله ، اندوتلیوم عروقی می تواند ساختار خود را حفظ کند. با افزایش تعداد سلولهای فوم ، آسیب اندوتلیال ایجاد می شود. آسیب به فعال شدن پلاکت کمک می کند. در نتیجه ، آنها ترمبوکسان ترشح می کنند ، که باعث تجمع پلاکت ها می شود ، و همچنین تولید فاکتور رشد مشتق از پلاکت را آغاز می کنند ، که باعث تقویت سلول های عضله صاف می شود. دومی از قسمت داخلی به لایه داخلی دیواره شریانی مهاجرت می کند و به این ترتیب در رشد پلاک نقش دارد. علاوه بر این ، پلاک ها با بافت فیبری جوانه می زنند ، سلول های زیر غشای فیبری نکروتیک هستند و کلسترول در فضای بین سلولی ذخیره می شود. در آخرین مراحل توسعه ، پلاک با نمک های کلسیم آغشته شده و بسیار متراکم می شود. در ناحیه پلاک ، لخته های خونی اغلب تشکیل می شوند ، لومن رگ را مسدود می کنند که منجر به نارسایی گردش خون حاد در محل بافت مربوطه و ایجاد حمله قلبی می شود. تنظیم سنتز کلسترول - آنزیم اصلی آن (HMG-CoA ردوکتاز) به روش های مختلفی انجام می شود. فسفوریلاسیون / دفسفوریلاسیون ردوکتاز HMG. با افزایش نسبت انسولین / گلوکاگون ، این آنزیم فسفریله می شود و به حالت فعال منتقل می شود. عمل انسولین از طریق 2 آنزیم انجام می شود. HMG-CoA ردوکتاز کیناز فسفاتاز ، که کیناز را به حالت دفسفریله غیرفعال تبدیل می کند: فسفوتاز HMG-CoA ردوکتاز با تبدیل آن به حالت فعال دفسفریله شده. نتیجه این واکنشها تشکیل یک فرم فعال دفسفریله شده از HMG-CoA ردوکتاز است. در نتیجه ، در دوره جذب ، کلسترول افزایش می یابد. در این دوره ، در دسترس بودن بستر اولیه برای سنتز کلسترول - استیل - CoA افزایش می یابد (در نتیجه خوردن مواد غذایی حاوی کربوهیدرات ها و چربی ها ، از آنجا که استیل CoA استیل در هنگام تجزیه گلوکز و اسیدهای چرب تشکیل می شود). در حالت پس از جذب ، گلوکاگون از طریق پروتئین ژناز A فسفوریلاسیون HMG - CoA - ردوکتاز را تحریک می کند ، و آن را به حالت غیرفعال تبدیل می کند. این عمل با این واقعیت تقویت می شود که در همان زمان گلوکاگون باعث تحریک فسفوریلاسیون و غیرفعال کردن فسفوتاز ردوکتاز HMG-CoA می شود ، بنابراین ردوکتاز HMG-CoA را در حالت غیرفعال فسفریله نگه می دارد. در نتیجه ، سنتز کلسترول در دوره پس از جذب و در طول روزه دار با سنتز درون زا مهار می شود. اگر میزان کلسترول موجود در مواد غذایی به 2٪ برسد ، در نتیجه سنتز کلسترول درون زا به شدت کاهش می یابد. اما قطع کامل سنتز کلسترول رخ نمی دهد. میزان مهار بیوسنتز کلسترول تحت تأثیر کلسترول ناشی از غذا در افراد متفاوت است. این نشان دهنده فردیت فرآیندهای تشکیل کلسترول است. با کاهش شدت سنتز کلسترول می توان غلظت آن را در خون کاهش داد. اگر تعادل بین دریافت کلسترول با مواد غذایی و سنتز آن در بدن از یک طرف و دفع اسیدهای صفراوی و کلسترول از طرف دیگر شکسته شود ، غلظت کلسترول در بافت ها و خون تغییر می کند. جدی ترین عواقب با افزایش غلظت کلسترول خون (هایپر کلسترول خون) همراه است ، در حالی که احتمال ابتلا به آترواسکلروز و کللیتیزیس افزایش می یابد. هایپرکلسترولمی خانوادگی (HCS) - این شکل بسیار متداول است - در حدود 200 بیمار در حدود 1 بیمار. نقص ارثی در HCS یک تخلف در جذب LDL توسط سلول ها است و بنابراین کاهش میزان کاتابولیسم LDL است. در نتیجه ، غلظت LDL در خون بالا می رود ، همچنین کلسترول از آنجا که مقدار زیادی از آن در LDL وجود دارد. بنابراین ، با HCS ، رسوب کلسترول در بافت ها ، به ویژه در پوست (زانتیوما) ، در دیواره شریان ها مشخص است. مهار سنتز ردوکتاز HMG-CoA محصول نهایی مسیر متابولیک کلسترول. این میزان رونویسی از ژن HMG-CoA ردوکتاز را کاهش می دهد ، بنابراین سنتز خاص خود را مهار می کند. کبد به طور فعال اسیدهای صفراوی را از کلسترول تولید می کند ، بنابراین اسیدهای صفراوی فعالیت ژن HMG-CoA ردوکتاز را مهار می کنند. از آنجا که ردوکتاز HMG-CoA پس از سنتز حدود 3 وجود دارد ، مهار سنتز این آنزیم کلسترول یک تنظیم مؤثر است.
|